�@���̃y�[�W�������������������̃o�[�W�����������p�������B�u���E�U�[�ł͌��ɂ����̂ł�������͑N���ł��B
�����̐i���_
�@�����̐i���_�̍��Z���x���̐����ł��B�������A�����̐i���_�͂�������Ȋw�I�E�̌n�I�Ƃ͌�����f�ГI�m���Ɋ�Â����������̐��_�Ɉˑ����Ă���B���̂��߂����̐��������ᖡ����c�_�ɍS�D��������S�̓I�ȗ���𗝉����邱�Ƃ���B�܂�i���_�̗����Ƃ͍��܂łɒ~�ς���Ă����l�X�ȉȊw�I�m���ƏƂ炵���킹�čł��m���炵�����J�j�Y����T���o����ƁB
�P�D�i���_�̕ϑJ
�i�P�j�V�ϒn�ِ��i�L���r�G�j
�@�u�V�ϒn�ق��n�������ʂ��Ċ��x���J��Ԃ���A���̂��тɑO�̎���̐����Q�͖w�ǎ��ł��A�n���̕Ћ��Ɏc�����������Q���V���ɍL�����z�����v�Ƃ������B�L���r�G�̓p���~�n�ɉ����锒���I�Ƒ�O�I�̒n�w���̉��̈Ⴂ���Ƃ����B�ނ͐��E�e�n�̉��ΐҒ�(������)���������������Ɣ�r���ċL�ڕ��ނ��A��r��U�w��ʂ��ČÐ����w�̊�b���m�������B
�@
�i�Q�j�p�s�p���i���}���N 1809�N�u�����N�w�v�A1815�N�u���Ғœ������v�j
�@�u���ɓK�����튯�͗ǂ��g�p����邩�甭�B���A�g�p����Ȃ��튯�͑މ�����B�����ς��Δ��B����튯���قȂ肻�ꂼ��̊��ɉ����Đ����͐i�����Ă����v�Ƃ������B
�@�����͌X�ɑn������s�ςł���ƌ����̂��펯�ł���������ɁA�����͕s�ςł͂Ȃ����ƂƂ��ɕω��������̂ł���ƁA�͂��߂Ă܂Ƃ܂�̂���̌n�I�Ȑi���_�\�����B�܂胉�}���N�͑Q�i�i�������߂ď������B�������l���`���̈�`���l���Ă����_�ɉ��Č���Ă����B
�@
�i�R�j���R�I����i�_�[�E�B��1869�N�u��̋N���v�j
�r�[�O�����̍q�C�@1831�N�i22�ˁj�`1836�N �w�r�[�O�����q�C�L�x�i1845�N�j
���C�G���́w�n���w�����x�i1830�N�j����傫�ȉe�����āA�q�C���̊ώ@�����u��͕ψق���A�����͐i������v�ƌ������z���B���������̒i�K�ł͕ψق̌������v�����Ă��Ȃ��B
�}���T�X�́w�l���_�x�i1798�N�j�����u���R�I���v�̒��z��B
�}���T�X�͐H�Ƃ͎Z�p�����I�ɂ����������Ȃ��̂ɁA�l���͊����I�ɑ�������X�������̂ŁA���R�̂܂܂ł͉ߏ�l���ɂ��H�ƕs���͔������Ȃ��Ƃ��A�l���𐧌����邽�߂ɗl�X�ȗv�����������Ƃ�_�����B
�ƒ{�A�͔|�A���̋N���ׁA�l�דI�I���ɕt���Č�������B
1859�N�i50�ˁj�w��̋N���x���A���R�����ɂ��K�Ґ����̌����\�B
�@�u�����̊ԂɋN�������ψق́A���ɓK�������̂��K�Ґ����ɂ�萶�������ɑł������Ďc�����A�����łȂ����̂��ŖS���Ă������R�I���ݏo���A���̎��R�I������S�̂�ψق����Ă����v�ƌ������B
1871�N�u�l�Ԃ̗R���v
�s���R�I���̗�t
- �I�I�V���t���G�_�V���N���H�ƈÉ�
�@���ȏ��ȂǂɕK���Љ��Ă����Ȃ̂ł����A���̐��������ɂ킩�ɐM����B�H�Ɖ��ɂ��A�̊��ɐ����锒���ۂ��n�ߗނ������ɂ�莀�ł��Ċ��������ۂ��Ȃ��āA���R�I���ɂ��É��^�̉�̊������������ƌ����Ă���B
�@���̃��J�j�Y���ɑ��ėl�X�Ȕᔻ�����邪�A�����̔ᔻ�҂��f����_�����I���˂Ă��Ȃ��B��������R�I���ɂ��ψق�����ɌŒ肵����ƍl���邩�炨�������̂��B���݂ł����Ì^���ÐF�^�����݂��Ă���̂�����B�����炱��͂S�D�ŏq�ׂ�ψق������Ď���ɌŒ肳�ꂽ���ʎ��R�I�𐫁i�K�Ґ����̓x����)���ω�������ɂ����Ȃ��B
- ����Ԍ����n�����̕��z�ƃ}�����A�����n�т̏d��
1954�N�@Anthony C. Allison. ����Ԍ����`���ƃ}�����A�ϐ��Ƃ̊֘A�����ؖ�
- �����̋[��
- �_��ɑ��鍩���̒�R��
- �R�������ɑ�����ر�̒�R��
�s�_�[�E�B���̌��сt
- ����Ȏ����������Đi���̏؋����������B
�@���̋L�^�A��̒n���I���z�A��r��U�w�A�����w�A�ƒ{�����ꂽ�����̕ψقȂǂɂ��đ�R�̏؋����W�߂Ď������B
�@�����A���Z�ŏK���i���̏؋��Ƃ��ẮA�����튯�������튯�A���Պ튯�A�K�����U���K���W���A�̔����͌n�������������A�n���I�Ȋu���ɔ����ώ�̕��z�A��Ӄ��J�j�Y���̕ϑJ�A�l�X�ȃ^���p�N�����`�q��ł����q�I�ȕψ��A���X������B
�@�������A�����̏؋��́A�����͑Q�i�I�ɏ������ω����Ă���A�l�X�ȕ����֕ω����Ă����ƌ������Ƃ𗝉������Ă���邪�A�Ȃ����̗l�ɕω����Ă����̂��A�ǂ̕����֕ω����Ă����̂��Ȃǂ̍��{�I�Ȗ₢�����̓����Ƃ͂Ȃ��Ă��Ȃ��B
- �i����Ҷƽ�тƂ��Ĕ��ɗL�͂Ȉ�̐����o�����B
�@���R�̎������A���Ȃ��Ƃ͂P�D�ŏW�ς��ꂽ�m������i���̃��J�j�Y����a���o�����ł���B�_�[�E�B�����a���o�������_�����R�I����ł���B
�s�_�[�E�B���̎��R�I����̌��E�ƌ��ׁt
- �P�D��̕����������o���Ȃ������B
- �@���Ƃ͐������ނ̊�{�P�ʁB�݂��Ɍ�z���Đ��B�\�͂�����q�����c�����Ƃ��ł���A�����͓���ƌ��Ȃ���A���Ƃ��q�����ł��Ă������ɐ��B�\�͂�������Έَ�ƌ��Ȃ����B�܂��Ƃ݂͌��Ɍ��G�����Ƃ��A�ɐB�\�͂̂���q�����c����W�c�̃O���[�v�̎��ł���B
�@���̂悤�ɐ��B�I�Ɍ��G�ł��Ȃ��O���[�v�ɕ�����Ă������J�j�Y�������܂������ł��Ȃ������B�@�@
- �R�D���R�I����ł͐����ł��Ȃ��ψق�����B
- �@�悭���������ɏo�����̂́A������̂ɂƂĂ��s���R�Ȋ튯�ƂȂ��Ă��܂����A�}�����X�̉�A�A�C�������h�I�I�c�m�W�J�̊p��A�˂�����Ȃ����Ă��܂����I�E���K�C�̊k�A�A�����i�C�g�̖D�����Ȃǂł��邪�A�����Ă�����ł��̕ω����L�����Ǝv���Ȃ��悤�ȕω��͎��R�E�ɂ͖����ɂ���B
- �R�D�ψق̌���������Ȃ������B
- �@�`���̈�`���i���`�q�̎��̂����炩�ɂȂ�̂��Q�O���I�ɓ����Ă����ł���B�_�[�E�B���̓��}���N�Ɠ����l�Ɋl���`������`����ƍl���Ă����悤���B
�@
�P�D�̌��ׂ����������̂Ƃ���
�i�S�j�u�����i1868�N�`�j
�u���R�I�������A����n��ɐ������u������邱�Ƃ��A�i���ɑ傫�ȍ�p���y�ڂ��v�ƌ������B
���[�O�i�[�i1868�N�j�n���I�u��
�@�@�@�@�@�@�@�@�@�@�@�嗤�̈ړ��A�����̐��v�◲�N
�@�@�@�@�@�@�@�@�@�@�@�C��̕ω��A�������A�X�͂̔��B�Ɉ���n���I���f
�@�@�@�@�@�@�@�@�@�@�@�C���A���ɂ��Ǔ��ւ̊u��
���}�[�j�Y�i1885�N�j���B�I�u��
�@�@�@�@�@�@�@�@�@�@�@���Ԋw�I��
�@�@�@�@�@�@�@�@�@�@�@�s���w�I��
�@�@�@�@�@�@�@�@�@�@�@���݂킯
�@
�Q�D�̌��ׂ����������̂Ƃ���
�i�T�j����i�����i��ϰ 1885�N�j
�u�����̐i���́A���Ƃ͖��W�ɁA�����̓��I�v���ɂ���Ĉ������i�ށB�v�Ƃ������B
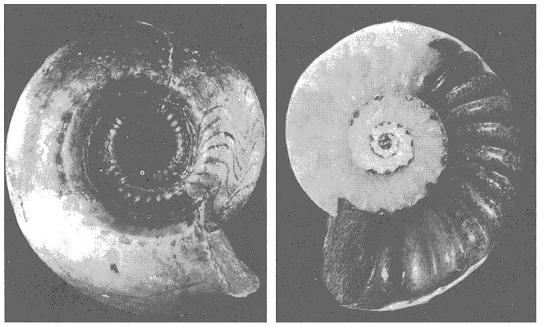
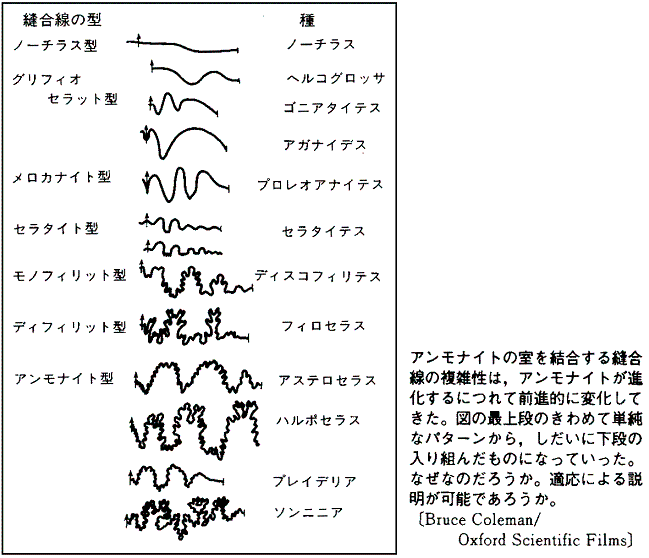
�@���ȏ��ł́A�s�K�v�ɋ��剻�����}�����X�̉��A�C�������h�I�I�c�m�W�J�̊p�A�s�K���ɕό`�����I�E���K�C�̊k��A�����i�C�g�̖D�����A����ƂƂ��ɑ�^�����Ă������n��ۂ̐i�������̗�Ƃ��ċ�������B
�@�������A�����ł͗Y�����ɔh��Œ������H��������傫�Ȋp�������́A��������Y�������I�Ԃ��Ƃɂ���Đi�����Ă����Ƃ������Ă���i���I�I���������j�B�������A�Ȃ������ɕs���ȓ��������Y�������I�Ԃ̂��A�܂������I���ʐ����c�闝�R�����炩�ł͂Ȃ��B

�@
�R�D�̌��ׂ����������̂Ƃ���
�i�U�j�ˑR�ψِ��i�ޥ�ذ� 1901�N�j
�@�I�I�}�c���C�O�T�̓ˑR�ψق̌�������o�������w���ŁA�u�����̐V������́A�������̓����̒��ɁA�ˑR�ψقɂ��啝�ȕψق��������̂��ˑR�����Ă���v�Ƃ�����B
�@�h�E�t���[�X��1886�N����T���{�ȏ�̃I�I�}�c���C�O�T�̖쐶�W�c�ׂĈ�`���̕ψَ킪�o�����邱�Ƃ������B���̂��Ƃ���A�ނ͐V������͎��R�����̍�p�̉��ŏ��X�Ɍ`�������̂ł͂Ȃ��A��`�����̓˔��I�ω��ɂ���Ĉꑫ��тɕω�����Ƃ����̂ł���B
�@�������ނ��������ψَ�̑����́A�����̈Ӗ��ł̈�`�q�ˑR�ψقł͂Ȃ��ِ��̂�{���̂₻�̑��̌��G�ُ�ɂ���Đ��������̂������B����̂ɋ}���Ȍ`���ψق������炵���̂��B
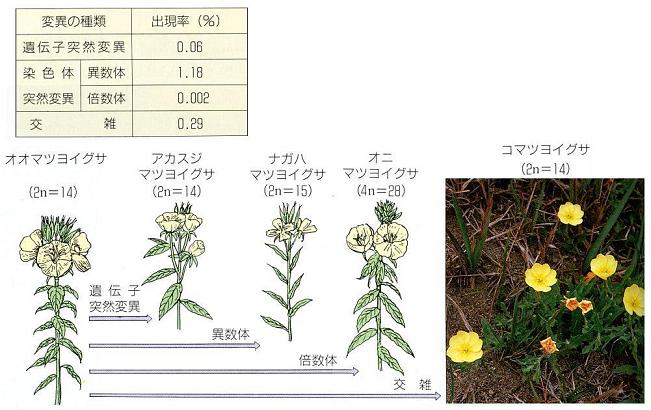
�@
�Ȃǂ��_�[�E�B���Ȍ�ɏo�Ă���B
�@���ɂP�X�O�O�N��O���̈�`�w�̋}���Ȑi���i���Ƀ��[�K����h�̃V���E�W���E�o�G��`�w�j�ɂ���āA��`�q�ˑR�ψق̂����ɂ͕\���^�̔��ɏ����Ȃ��̂����邱�Ƃ�����A��`�̗L�l�ƓˑR�ψق̎��̂����炩�ɂȂ��Ă���B
�@��`�w�̐V�����m���Ǝ��R�I����́A�₪�đ����i�����Ƃ����`�œ�������Ă����B
�@
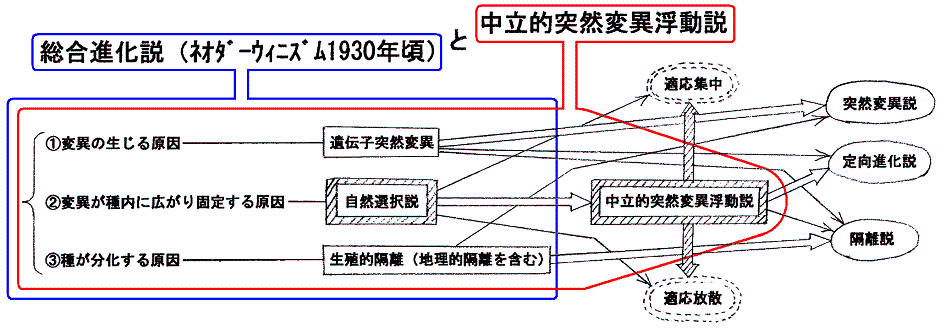
�Q�D�����i�����i�l�I�_�[�E�B�j�Y���@1930�N���j
�@�����i�����Ƃ́A�ψق̌������ˑR�ψ��ŁA�ψق�����ɍL�܂�Œ肵�čs�����������R�I���ŁA�����Ĉ�̎킪�l�X�Ȏ�ɕ������Ă����������n���I�E���B�I�u���Ő���������̂ł���B
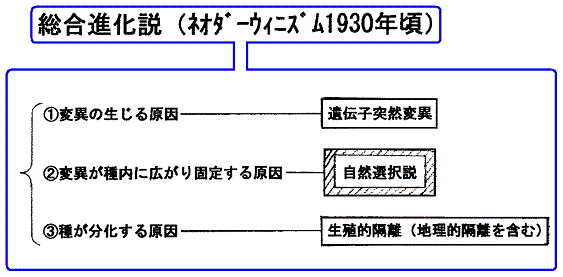
�@����`������O���[�v���̂���̂ɓˑR�ψق�������Ƃ���͎��R�ɂ��̃O���[�v�S�̂ɍL����Ǝv����������Ȃ����A�O���[�v���`������̐��������āA�O���[�v���̎��R�Ȍ��G���ۏႳ��Ă���ꍇ�ɂ͂����͂Ȃ�Ȃ��B������n�[�f�B�E���C���x���O�̖@���i1908�N�j�Ƃ����B
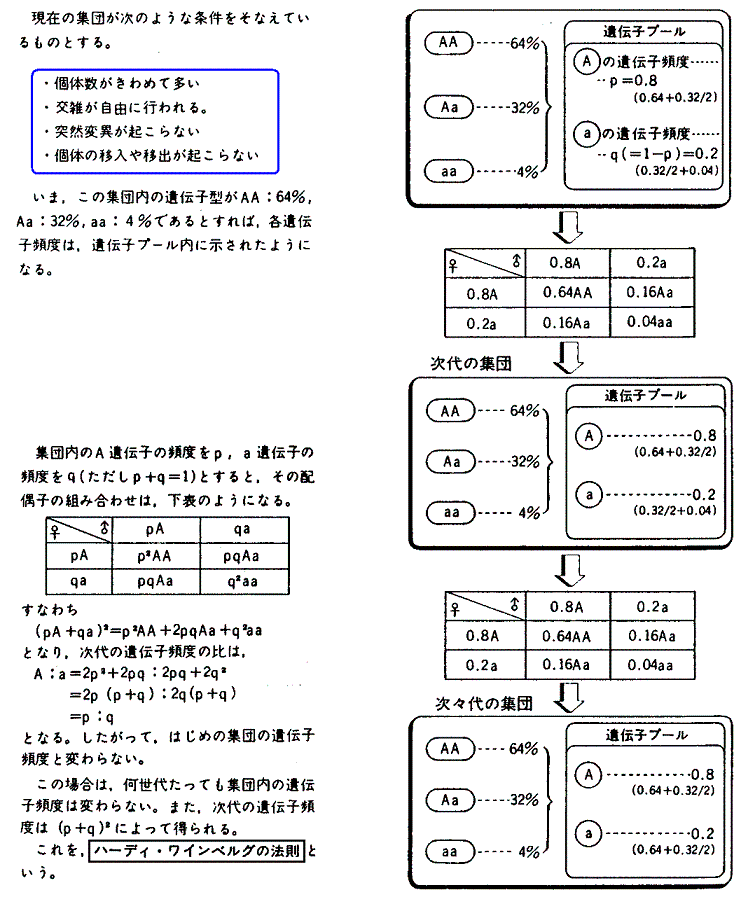
�@���̂Ƃ����R�I���������ƕψق��W�c���ɍL�܂�Œ艻���Ă����B�܂����R�I���̖��������ɂ߁A�ψق̌�������̕����Ƃ̊W�m�������̂������i�����ł���B
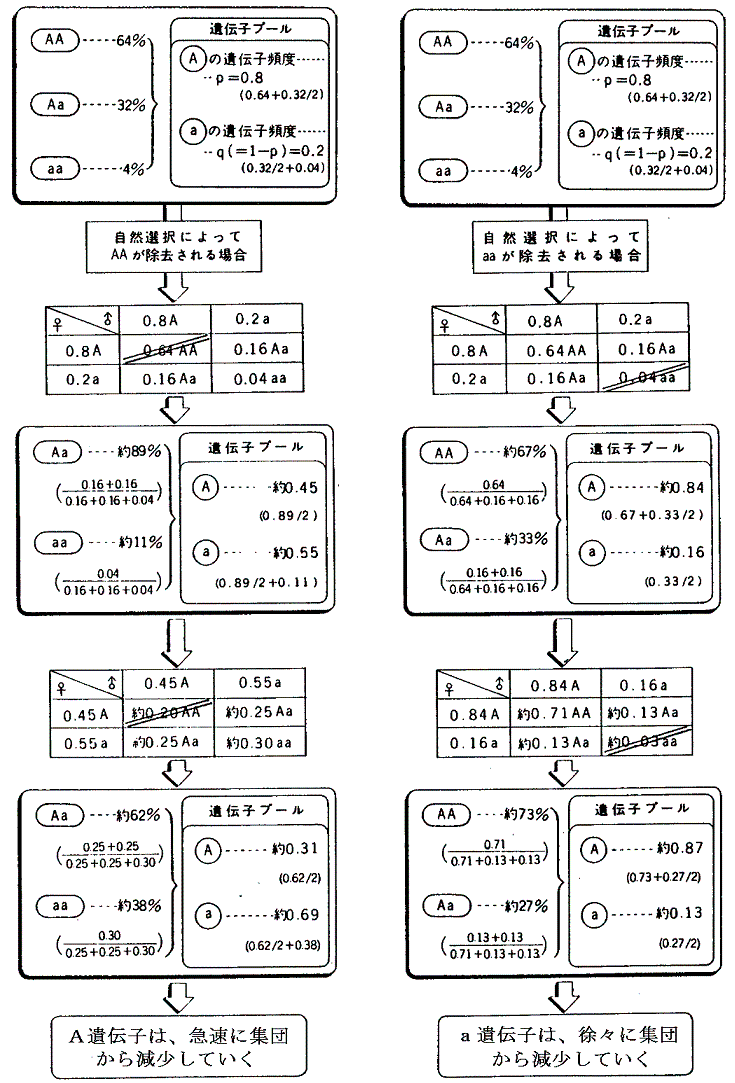
�i���_�Ƃ��Ă̑̍ق͂��Ȃ萮���Ă������A����ł��ȉ��̗l�Ȗ��_�����݂����B
�s�����i�����̖��_�t
- ���ʂɐ�����ˑR�ψق͂��̂قƂ�ǂ��A���K�͂Ȃ��̂ł��ꂪ����������ƌ����āA���ꂪ�����Ɏ��R�����ɑ��ėL���ɂȂ�����s���ɂȂ����肷����̂ł͂Ȃ��B����ɓˑR�ψق̑命���͓łɂ���ɂ��Ȃ�Ȃ������I�Ȃ��̂��L�Q�Ȃ��̂ł���B
- ���A�̒��ɏZ�ލ����̖ڂ�F�f���މ����Ă����悤�ȁA���̓ˑR�ψقɂ��튯�̑މ�������ł��Ȃ��B���A�̈Èłł͖ڂ������Ȃ����Ƃ͗L���Ȃ��Ƃł͂Ȃ��B����Ȃ̂ɈÍ��̊��ɐi�o���������͒Z���Ԃ̐�����ł��̂�����\�͂������Ă����B�l�ނ��T�疜�N�O�Ɏ������r�^�~���b����������\�͂�A�k�o�̑މ��Ȃǂ����̗l�Ȃ��̂��Ƃ�����B���������������މ��̕��q�_�I�ȏ؋��͑�R����B
- ���B�r���̊튯�͐����ɗL���Ƃ͌����Ȃ��B�ނ���ז��ɂȂ���̂̂ق��������B
- �ߓx�ɔ��B�������ɗL���Ƃ͍l�����Ȃ��튯��������������B
- ��ʂɑ�W�c�قǎ���̕ψق͏������B
- �i�����x�͏��W�c�قǑ����B�i��@����ߺ�����j
- �i���������I�ɐi�����i��i���j��i������r�I�������i�����i���i���j�����邱�Ƃ�����ł��Ȃ��B
���Ð��㒆���ɍ��������߂ď㗤���čL��ȑ�n��V�������Ƃ��Ċl����������
���Ð��㖖���璆���㏉���ɂ����ĐҒœ����������ށA��ނւƐi�����Ċ��S�ɐ������E����Ɨ����čL��ȓ������ɐi�o��������
�������̐�łɂ���ԂɂȂ����ɐi�o�����M���ނ������I�ɐi�������V���㏉��
�����̖�������C�ɉ�������l���������ɏq�ׂ钆���I�ˑR�ψٕ������ł���B
�@
�R�D�����I�ˑR�ψٕ������i�ؑ�����1968�N�A�L���O�A�W���[�N�X�@1969�N�j
�@1968�N�ؑ������i�ĵ�j�ɂ���čŏ��ɏ�����ꂽ���B�����I�ˑR�ψٕ������̓��e���̂��̂͒���i���������A���q��`�w���W�c��`�w�i���w�I�A���v�w�I�Ɉ�`����������j�̔��W��҂��č��̐��̓W�J���\�ɂȂ����B
�@���Ɏ���`������W�c���������ꍇ�A���v�I�Ȃ�炬�̂��߂Ɏ��R�I���������Ȃ��Ă��ψق��W�c���ɍL����Œ肵�Ă����i���C�g���ʁi1931�N�j�j���Ƃ��N����A���ꂪ���̐��̒��S���Ȃ��B
�s���̐����x������l�X�ȋ�̓I�m���t
�@�}�E�X�Ŕ������ꂽ�w���O���r���̋U��`�q�̔����ƋU��`�q�̐i�����x�̌����Ȃǂ���n�܂�A�����ł͖c��Ȑ��̕��q�I�ȏ؋����W�܂����B���݂͂����𑍍��I�ɘ��Ղ��Ă܂Ƃ߂�i�K�ɒB���Ă��Ȃ��B��`�q�̉�Ǎ�Ƃɂ��A���X�ƕ��q�I�ȐV�������������炩�ɂȂ��Ă��鎞��ł���B�����̍e�́A�����������Ԃ��o�đS�̓I�ȑ��������Ă������Ďn�߂Ă��܂��܂Ƃ߂���Ǝv����̂ŏڍׂ͏ȗ��B�i�ŋ߂̕��q��`�w�̐��ʂ́@�{�c�����́@http://www.brh.co.jp/katari/shinka/�@�������������j
�@������ɂ��Ă��A���q�����w�I�ȃA�v���[�`���\�ɂȂ��āA�n�߂Đi���_�͘_���I�Ɍ��ł���Ȋw�I���_�ɂȂ����B
�@
�s���̐����w�������i���̃V�i���I�t
�@���̐��̖{���́A��`�q�̏�ɕ��q�I�ȕψق͎l�Z�����N�����Ă���A������\�ȕ����֕ω����Ă��鎖�ł���B���̈�`�q�ψقɔ����\���^�̕ω��͑����̏ꍇ�L�Q���A�����I�Ȃ��̂ł���B�L�Q�ȏꍇ���̎q���͐����c��Ȃ��̂ŁA�����ɏW�c�̒������菜�����B�����瑽���̏ꍇ�����I�ȕψق݂̂��W�c�̒��ɌŒ艻���ꎟ����Ɏp����Ă����B
�@���̂Ƃ����v�I�ɏW�c���������قǁA���v�I�Ȃ�炬�̂��߂ɁA�����̕ψق͏W�c���ɐ����c��Ղ��Œ艻����₷���B���̂Ƃ��X�̕ω��͏��������ω��̕����͗l�X�ȕ����ɃA�g�����_���ɋN����B�Ⴆ�Ηl�X�Ȉ�`�q�������I�Ɋ֗^���鐶���̑傫���ɂ��Ă���^���̕����ւ��A���^���̕����ւ�����������ɋN����B���ׂĂ̕����ɕω����Ă����̂ł���B
�@���̂Ƃ��A�Ð��㖖�⒆���㖖�ɋN�������l�Ȏ��R���̌��ςɂ��A�����̐����킪��ʂɐ�ł��čL������Ԋw�I���i�G�R���W�J���j�b�`�F�j���ɂȂ����ꍇ�A���ꂼ��̊��ɓK�����ē��ꉻ���Ă��Ȃ����n�I�Ȏ킩��o�����āA���ꂼ��̊��ɓK�������l�X�Ȏ킪�����I�ɐ��܂�Ă��āA�����̃j�b�`�F���߂Ȃ����B
�@���̌��ςɑς��Đ����c��̂́A�����̏ꍇ����܂ł̊��ɓK�������ꉻ���Đ��Ԍn�i�H���A���j�̍ŏ�ʂ��߂Ă������B����������ł͂Ȃ��āA���̕Ћ��ɂЂ�����Ɛ����ē��ꉻ���Ă��Ȃ��i���ꉻ���Ă��Ȃ�������K���\�͂������j���n�I�Ȑ����ł���B�������������n�I�Ȑ����킩��K�����U���n�܂�B
�@���̂Ƃ��A�K�����U�Ƃ͎��R�I���ɂ����ɓK�������킪�����c�����̂ł͂Ȃ��āA�����I�ˑR�ψق̐ςݏd�˂ɂ�肠��������ɐi�����Ă����l�X�ȕψَ킪�A���ꂼ��̐������ɓK�����Ƃ���i�o���čs�����ۂł���B�܂�A���ɂ�鎩�R�����Ƃ��������A�����I�ˑR�ψق̐ςݏd�˂ɂ��ω����Ă����l�X�Ȏ킪�A��̊��ɉ�����K�����Ő����������J��L���A���̃j�b�`�F�ɍł��K�������킪�����c���Ă����B
�@�l�X�Ȋ��ɐi�o���Ă������Ƃ́A�K�R�I�ɒn���I�Ȋu���A�����K���ɂ�鎞�ԓI�E�ꏊ�I�Ȋu���ݏo���B�����̊u���ɂ�萶�B�I�Ȋu������������B���B�I�Ȋu�����m��������Ԃł���ɕψق��ςݏd�Ȃ��Ă����ƁA�₪�Ă����̕ψَ�̊ԂŎ��Y����z���Ă����B�\�͂̂���q�����̂����Ȃ��Ȃ�B�܂��̕��������������ƂɂȂ�B�������ĕʁX�̎�ƂȂ�A�܂��܂����ꂼ�ꂪ����ȕ����i�����Ă����B
�@�����āA�����̏���ȕω����i�݂����A���ꂼ��̊��ւ̓K�����Ɍ��܂ł����ނƁA�����̏ꍇ���Ԋw�I���̕ω��ɑ���K�����������A�����킸���Ȑ��Ԋw�I�Ȏ��R���̕ω��Ő�ł��Ă��܂��B�܂��n���I�ȕϓ��ɔ��Ȃ��Ċg�U�E�������Ă����ʎ�̓��A���Ƃ̐��������ɕ����Đ�ł��Ă��܂��B
�@�قƂ�ǂ̎킪�l�X�ȏ̐i���̑��H�ɓ���Ō�ɂ͐�ł��Ă����B�܂�l�Z���������̎�͐�ł��Ă���A�V�����킪����Ɠ���ւ��ɂ�����Ă���B�������A���ւ̓K�����ɗD��Ă���A�����N���ɓn���Đ������тĂ����A�����Ă��鉻�Ƃ�����悤�Ȑ����������B
�@�����̐��E�ł́A�H���A������ϓ��̒��ŁA���R���x�z����l�X�ȗv���ɂ��A�a������q���̂����ꕔ�݂̂������c�莟����̎q�����̂������Ƃ��ł��邪�A��̃��x���̑����Ɛ�łɉ����Ă��A���̑����͋��R�Ɏx�z�����B
�@
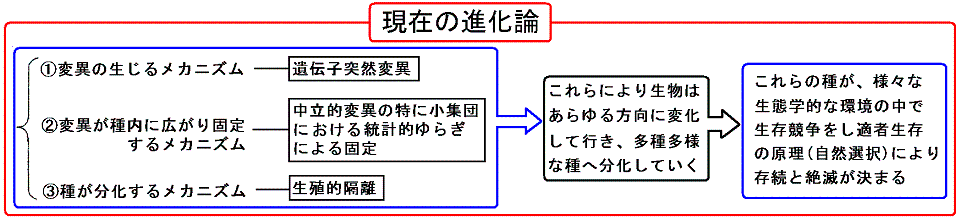
�S�D���݂̐i���_
�@�����Œ��ӂ��Ȃ���Ȃ�Ȃ��̂́A�����I�ˑR�ψق̓��v�w�I�����ɂ�����Œ肪�ψفi�i���j�̎�v�������A���̂悤�ɂ��ĕψفi�i���j���Ă�����������Ƃ��Đ����c���Ă����邩�ǂ����́A���Ԋw�I���ւ̓K�����ɂ���B�܂�A���R�I����ŏq�ׂ鏊�̓K�Ґ����̌����ł���B���������̓����Ƃ���́A�l�X�Ȑ������ω����A�܂��킪�������Ă��������Ƃ��Ăł͂Ȃ��B�����I�ˑR�ψق̓��v�w�I�����ɂ�����Œ�ɂ�蔭�W���Ă����l�X�Ȑ����킪��Ƃ��đ������Ă����邩�ǂ�����������郁�J�j�Y�������R�I���ł���B�����̔���������ŏI�i�K�œ����Ă���B�����������Ӗ��ŁA�_�[�E�B���̏��������R�I����͏����Ă����ǂ��납�܂��܂��d�v���𑝂��Ă���B
�@���̂悤�ɉ��߂���A���܂Ńo���o���ɋc�_����Ă�������i�����A�u�����A���B�I�u���A���I�I����A���ݕ������A�K���W���A�K�����U�A��i���A���i�����X�̒f�Ђ��A�i���_�̑S�̑��̒��ŁA���ꂼ��̏ꏊ�Ƀs�b�^���Ǝ��܂���A���ׂĂ�I�ɗ����ł���B
�@
�i�P�j�����O���[�v�i��j���ω��i�i���j���A�܂���̎킪���l�Ȏ�֕������čs�����J�j�Y��
�@�ψق̐����郁�J�j�Y���@���@��`�q�ˑR�ψ�
�@�����A��`�q�ɐ�����ˑR�ψق́@���ː��A���w�����A���O���E�w�����̕����I�v���A��ٽ�����@���������ƂȂ��Đ����邱�Ƃ������Ă���B�����āA�����̗v���͐��B�זE�̈�`�q�Ɏl�Z�����e����^���Ă���A��`�q�́A�l�Z�����������ω����Ă���B�����̏ꍇ�����̏��͕��q�I�ɒ����ɏC������邪�A��Ɏ��̐���Ɏp�����B
�@���̂悤�ɂ��Đ������`�q��̕��q�I�ȕω��́A�������ɍ��o�����^���p�N���̐�����ω�������B���̕ω��͂قƂ�ǂ��L�Q�Ȃ��̂������I�Ȃ��̂ł���B�L�Q�ȕψق͎̌̂��ł��Ē����ɁA���̈�`�q�ω��͐����W�c�����菜����邪�A�����I�Ȃ��̂́A�̃��x���Ő����c��B
�@���Z�����ŏK���悤�ɁA���B�זE���`�����錸������́A��`�q����F�̏�Ɋ������A�����A�����̂Ɩa���̂ɂ�镪�������ɕ��G�Ńf���P�[�g�Ȍ��ۂł���B���̂��߁A�����ΐ��F�̂̕����╪���Ɍ�肪������B���̌��ʂƂ��Đ��ݏo����鐶�B�זE�i���q�E���q�j����̈�`�q(���F�́j�ɂ��t�ʁA�d���A�����A�]���A�Ȃǂُ̈킪������B�܂������̗Z���ɂ���Đ��F�̂̒����̕ω��␔�̕ω�����������A���������܂����������F�̐��̐����{������悤�Ȍ��ۂ�������B
�@����炪�����̐��B�זE����ɋN����ƁA�����̏ꍇ�����̐���ɉ����Ēv���I���ׂƂȂ�B����͓����̑̂������̉ߒ��ŕ��G�Ŕ����Ȓ����̊�ɍ���ċ��邩��ł���B�������A��ɂ��̂܂܂Ő������ѓ���ψقƂȂ�ꍇ������B
�@�܂��A���ɋN����A�A���͓����ƈ���đ̂̂���͒P���ŁA����̍זE��������ςݏd�˂�悤�ɏW�����č\������Ă���̂ŁA���I�Ȍ`���ω����Ȃ�����������т�ꍇ������B��ʂɐA���͓����ɔ�r���āA�������������F�̃��x���̕ψقɂ��Ă̑ϐ��͍����B
�A�ψق�����ɍL����Œ肷�郁�J�j�Y���@���@�����I�ˑR�ψق̓��v�I�Ȃ�炬�ɂ�����ł̌Œ�
�@�@�̕ψقɂ���āA�̂̒��ɐ������т���`�q���x���̕ω��́A��ʂɂ͐�����J��Ԃ��ɂ�āA�W�c�̒����玸���Ă��܂��B���������ɂ́A���v�I��炬�̂��߂ɏW�c���\������̂̑S�̂ɍL����A��Ƃ��Ĉ�`�q���ω����Ă�������������B�����́A�܂��ɋ��R���x�z���铝�v�I�A�m���_�I�Ȍ��ۂł���B
�@���̂Ƃ��A���w�I�ɖ��炩�Ȃ悤�ɁA������������炬�ɔ����ĕψق��W�c���֊g�U���Œ艻����m���́A�W�c���\������̂̐������������̂قǑ傫���Ȃ�B�W�c���\������̂̐��Ƃ́A���̏W�c���̊e�̂��A���̏W�c���̑��̌̂Ɠ����Ȋm���Ō݂��Ɍ�z������ɂ���O���[�v�̌̐��̂��Ƃł���B
�@�܂萶�B�I�Ɋu�����ꂽ���W�c�ɕ������鎖���d�v�ɂȂ�B�W�c���������قǐi���̃X�s�[�h���������Ƃ́A���ۂ̐����E�̗l�X�ȗ�Ŏw�E����Ă���B�����āA���ۂ̏�������Ƃ���ŏ��W�c���͎�������Ă���B�n���I�Ɋg�U���邾���ŁA���̊e�X�͏��W�c�ɕ��������B�Ⴆ�A�����z���E�T�s�G���X�ł��A���̏Z�ޏꏊ���g�U���邱�ƂŁA�l�O���C�h�A�R�[�J�\�C�h�A�����S���C�h�A���X�ɕ�����Ă������B����ɓ����R�[�J�\�C�h�A�����S���C�h�ƌ����Ă����S�L������č����Ⴆ�Δ��̐F�E����E�̂����ς���Ă��邱�Ƃ́A����̍��𗷂��Ă݂�Β����ɗ����ł���B�܂�A���ł����Əڂ����q�ׂ�悤�ɁA��������@�œ��v�I��炬�̌Œ肪�N����₷�����W�c���������Ă����̂��B
�B�킪�������郁�J�j�Y���@���@���B�I�u��
�@���łɏq�ׂ��悤�ɁA�A�ŏq�ׂ����Ƃ������������ׂɂ͎킪���W�c�ɕ�������邱�Ƃ��K�v�ł���B���̂Ƃ���Ȃ��Ƃ݂͌��Ɍ�z�ł���E�ł��Ȃ��A�̐��B�I�Ȋu������������邱�Ƃł���B���B�I�Ȋu���̐����ɂ͗l�X�ȃ��J�j�Y�����l������B
�@�Ⴆ�A��m�v���[�g��̃z�b�g�X�|�b�g�ɐ�����ΎR���Ȃǂ̂悤�ɁA��C�̌Ǔ��Ƃ��Đ��������ɁA�l�X�ȗv���ŗ��ꒅ���������킪�����œƎ��̔��W������B���邢�́A�X�͊��ƊԕX���̌J��Ԃ��ɂ��C���ʂ̕ϓ��i�P�T�O�����z����j�ɔ����u�����ꂽ�藤�����ƂȂ铇�X��嗤�B���́A�v���[�g�e�N�g�j�N�X�ɔ����ړ��ɂ���ė����W�U����嗤�Ⓡ�ʁB
�@����ɁA�C��ϓ��ɔ��Ȃ�������A�X�т̏��łȂǂ̐A���E���̕ω��ɂ�鐶�Ԋw�I�ȕ����Ŏ�������鐶�B�I�u���B�܂��́A���Ԋw�I�Ȑ����l���̕��U�ɂ��A�ꏊ�I�ɂ͓����n��ɏZ�݂Ȃ���A������������ƒn��A���ƊC�A�ȂLjقȂ����������ɕ������邱�ƂŐ��B�I�Ɋu�������B�@���邢�́A�����̎��ԑт�����邱�Ƃɂ��A�����ꏊ�A�������ɍς݂Ȃ��琶�B�I�Ɋu�������B���X�l�X�ȏ��l������B���ݔ���ȗႪ�A���ۂɊώ@�������Ă���B
�@���̂悤�ɂ��āA��U���B�I�Ȋu�����m�������e�O���[�v�ł́A���ꂼ��̃O���[�v�ɌŗL�ȕψفi������m���_�I�ȕ����E���U�Ɉˑ�����j���~�ς��Ă����B�����ω����ςݏd�Ȃ�ƁA�����̃O���[�v�ԂŌ�z�������Ă��A�ɐB�\�͂̂���q�����c���Ȃ��Ȃ�A��Ƃ��Ă̕������m�����邱�ƂɂȂ�B�����Ȃ�ƁA���ω����Ēn���I�A���Ԋw�I�ȕ�������������Ă��A���͂�B�I�Ȋu�����������鎖�͂Ȃ��̂ŁA�킲�ƂɕʁX�̕ω����W�ς��Ă����B
�@
�i�Q�j���l�����A�ω����Ă����l�X�Ȏ킪�����E���W�E��ł��郁�J�j�Y���Ƃ��Ă̎��R�I��
�@�O���̇@�A�B�̃��J�j�Y���Ői�����Ă��������̗l�X�Ȓi�K�A�l�X�ȏœK�Ґ����̎��R�I���������A��Ƃ��Đ��S�E�������Ă����B���̂Ƃ��l�X�ȏɉ����郁�J�j�Y�����l������B
- �O�J���u���A�I�ɒP�זE���������זE���̔\�͂��l�������Ƃ��B�����̑̂̍����Ɋv���I�ȕ��@�đ����ʂ֔����I�ɐi�������B�G�f�B�A�J�������Q���o�[�W�F�X�����Q
- ��C��w���ɃI�]���w�̌`���Ƃ����v���I�Ȋ��ω����������Ƃ��B�����ɗ���Ƃ����S���V�����L��ȃj�b�`�F�����p�\�ɂȂ����B�C�����ŏ�L�@�A�B�̃��J�j�Y���ł���������֕ω����������̂�������ɓK����(������h���̕\�A�̂��x���t�������点��ۊǑ���)�@�\���l���������̂��A���Ƃ��čŏ��ɏ㗤���ʂ������V�����I�B
- �����ōŏ��ɏ㗤�ł����̂́A�����ł��łɗ���ł̐����ɕK�v�Ȍ`���i������h�������k�A�n����͂���邽�߂̑��j���l�����Ă����ߑ������ł���B�ނ�͓����ɂƂ��Ė����̍L��ȃj�b�`�F�ɔ����I�ɓK�����U���A��^�������B
- �̓��ƌł��k�Ɨr���A�r�����������l�����Ċ��S�ɐ������痤�ւ̐i�o���ʂ������Ғœ����B�ނ�͂��̗D�ꂽ�^���@�\�ɂ���āA��ɐi�o���Ă��������ނ��������Ă����A�����͏��^�����ĕʂȐ����c�����I�����邱�ƂɂȂ����B
- �Ð���ƒ�����̎���敪�̍����ƂȂ��K�͂Ȑ�ł������N�������A�ΎR���������n�k�ϓ��ɂ����ω��B
- �����㏉���ɂ́A����ōŏ��ɉh�����Ғœ����ł���P�|�ޓ������A���D�ꂽ�̌^�Ɖ^���@�\���������ށi�o�|�ށj�Ɉ�������Đ��ށB
- ���̗D�ꂽ�̌^�Ƒ�Ӌ@�\�ɂ��A������ɂ͋����ނ�������j�b�`�F�ɐi�o�����B
- �����㖖��覐̏Փ˂ɂ������ρB����ɂ���ł��������ނ̃j�b�`�F���A����܂ň�������ĂЂ�����Ɛ������Ă����M���ނ������I�ɐi�����Đ�߂Ȃ������B
- �p���Q�A�嗤�̕���ɂ��嗤�ړ��ɂ�蕪���Ǘ��������I�[�X�g�����A�嗤�ł́A�L�ܗނ�������j�b�`�F�ɐi�o�����B
- ���嗤�ł́A���D�ꂽ�玙�@�\�����L�ٔނ�����āA������j�b�`�F�ɐi�o�����B
- �l�ސi���̉ߒ��Ő�ł��Ă������q�g���̗l�X�Ȏ��B�N���}�j�����l�Ɉ�������Đ�ł����l�A���f���^�[���l�ȊO�ɂ��A�i���̑��H�ɓ���ŖS�����l�ނ̈���͑�R����ƍl�����Ă���B���u�X�g�X��{�C�Z�C�ł�����o�����g���v�X����A�W�������l��k�����l�Œm����z���E�G���N�g�X�������̂悤�Ȑ�Ŏ킾�ƍl�����Ă���B
- �X�͎���̊C�ʒቺ�ɂ�藤���Ōq���ꂽ��k�A�����J�嗤�ɐl�ނ��i�o�B�����Đl�ނɂ�葽���̑�^�M���ނ���ł����B
- �ߔN�ɂȂ��ă��[���b�p�l���������L�ٔނɂ��A���̐�������������Đ�ł��邱�ƂɂȂ����I�[�X�g�����A�̋�������j�b�`�F�̗L�ܗށB
- �X�y�C���l�̒��āA��đ嗤�ւ̐i�o�ɂ���Ď������܂ꂽ�V�R���Ȃǂ̕a���ۂɂ���ł̊�@�ɕm�������n�Z���B
- �l�ނ̑��B�ɂ���Đ���������Ȑ��̐�����̐�ŁB�Ⴆ�|�[���E�G���b�N�A�A���E�G���b�N�����u��ł̂䂭��(�����̑��l���Ɛl�ނ̊�@�j�v�V�j��1992�N���@�Ȃǂ�ǂ܂ꂽ���B�����ɂ͋����ׂ����̐�ł̗Ⴊ�L�ڂ���Ă���B
- ���j��̗l�X�ȕ����Љ�E�����̗����E���S���A���݂̍��Ƃ̕ϖe�����ǂ��̂悤�Ȃ��̂ł���B
�@���̂悤�����������ɉ�����K�Ґ����̌����Ɋ�Â����R�I���͗l�X�ȃ��x���E���J�j�Y���E�l���E�`�ԂŐ������B��X�̌Ð����w��Ȋw�I�Ȓm����������ɂꎩ�R�I���̓��e�̗����͐[�܂��Ă����B
�@
�i�R�j���݂̐��E�̂�����Ƃ���ŋ@�\���Ă���K�Ґ����̌����Ɋ�Â��ω�
�@�����͂₪�ėl�X�Ȓm����~���p������\�͂��l�������B���̂��߁A���̎Љ�̒��ɐ������Ȋw�I�Ȕ����A�Z�p�I�Ȕ����A�Љ�x�̉��ǁA�����l���̉��Ǔ���������Čp�����ꂾ�����B����͂��ꂼ��̐�����Ɉ�`�I�ɖ{�\�̌`�Œ~�ς���Ă����m���A�K���𗽉킵�Đ����̐����ɉe����^���n�߂��B
�@���ɐl�ނ��l�������A���s�ɂ���Ď��R�ɂȂ����O���̐i���ɂ���ĉ\�ɂȂ����Ί�i����j�̎g�p�A�̎g�p�B�܂��A�������s�ɂ�����ю���̍\�����ω����Ăł���悤�ɂȂ����l�X�ȉ����𑀂�\�͂ɂ�茾�t������ׂ��悤�ɂȂ������ƁB�l�X�ȏ���`�B���錾��̗��p�B���������Z�p�̔����ɂ����̒~�ςƓ`�B�B�R���s���[�^�̔����ƃC���^�[�l�b�g�v���ɂ�闘�p�ł�����̑�K�͉��ƌ������B���邢�͗��j�Ղ̔����ɂ��q�C�p�̐i���A�|�⑄��e�̔����A���C�@�ւ���R�@�ւ̔����A�d�C�̔����Ɨ��p���X�A�Љ�̐��Ƃ��Ă̋����̂̌`���A���Ƃɂ��E�Ƃ̍��x���A�����`�̊m���A���R�����Ɋ�Â����{��`�̌����A���X�Ȃǂ̂�����ω��������Ƃ��Ă̐������E�����E���S�̗������߂�B
�@�����̕ω��͂���������ɋN�����Ă���A���̕ω��̒��ł����ꕔ���K�Ґ����Ő����c�蔭�W���Ă����B����ɂ�荑�Ƃ����W�E�ŖS���A�l�X�ȎЉ������q���ĕω����Ă���B���ݎЉ�ł̗l�X�Ȋ�Ƃ̑��S��A�g�̉��ŗ��p����鏤�i���K�Ґ����̌����œ�������Ă����B
�@���̂悤�ɂ�����ω��ɓK�Ґ����̌����������Ă���Ƃ����F���ɗ��ƁA���߂ēK�҂Ƃ͉����A�����Ƃ͉����A�������i�����W����Ƃ͉����A�l�ԂɂƂ��čK���Ƃ͂Ȃɂ�������Ă���B�������A�����̖₢�ɐ����������邱�Ƃ͉i���ɖ������낤�B