丂偙偺儁乕僕傪報嶞偝傟傞曽偼偙偪傜偺僶乕僕儑儞傪偛棙梡壓偝偄丅僽儔僂僓乕偱偼尒偵偔偄偺偱偡偑報嶞偼慛柧偱偡丅
丂惗暔傪偳偺傛偆偵暘椶偡傞偺偐偼擄偟偄栤戣偱偡丅惗暔偺帠偑傛傝徻偟偔夝偭偰偔傞偵偮傟偰暘椶懱宯偼夵掶丒夵曄傪偔傝偐偊偟偰偒偨丅偟偐偟丄崱擔偱偼暘巕惗暔妛揑側堚揱巕夝愅偵傛傝恑壔偺摴嬝傪尩枾偵扝傞偙偲偑偱偒傞傛偆偵側偭偨丅偦偺偨傔壢妛揑崻嫆傪帩偭偰暘椶偱偒傞傛偆偵側傝丄宯摑庽偼傛傝怣棅偱偒傞傕偺偵側傝偮偮偁傞丅
丂崱擔丄惗暔偼丂尨妀惗暔偺乮尨妀惗暔奅乯丄恀妀惗暔偺乮尨惗惗暔奅丄怉暔奅丄嬠奅丄摦暔奅乯丂偺俆奅偵暘偗傞偺偑堦斒揑偱偁傞丅尨妀惗暔偲偼堚揱巕偑妀枌偵曪傑傟偰偄側偄尨巒揑側嵶朎偺惗暔偱偁傞丅恀妀惗暔偲偼堚揱巕偑妀枌偵曪傑傟偰崅搙偵婡擻揑偵廂擺偝傟偰偍傝丄嵶朎撪偵偼妀埲奜偵儈僩僐儞僪儕傾丄彫朎懱丄僑儖僕懱丄梩椢懱側偳偺崅搙側枌峔憿偺婍姱傪帩偮嵶朎偱偱偒偰偄傞惗暔偱偁傞丅崱擔丄偐偭偰怉暔奅偵偄傟傜傟偰偄偨憯椶傗嬠奅偺棏嬠椶丒擲嬠椶偼尨惗惗暔奅偵堏偝傟丄尨惗惗暔奅偼懡嵶朎惗暔傕娷傓傛偆偵側偭偨丅
丂惗暔偼丂奅仺栧仺峧仺栚仺壢仺懏仺庬丂偺弴斣偱師乆偵嵶暘偝傟偰偄傞丅偙傟偱懌傝側偄偲偒偼乽忋乿丄乽垷乿丄乽壓乿傪晅偗傞丅椺偊偽丂忋栚仺栚仺垷栚仺壓栚丂偺條偵丅
丂摦暔奅偼廬懏塰梴偺偨傔偵怘暔傪昁梫偲偡傞丅偦偺偨傔怘傋暔傪媮傔偰摦偒夞丄怘傋暔傪庢傝崬傒丄徚壔偡傞婡擻傪帩偨偹偽側傜側偐偭偨丅偮傑傝摦暔奅偼懱偺嶌傝曽偺夵椙偱恑壔偟偰偒偨丅
丂乮偙傟偼惗怋朄傪夵椙偡傞帠偱恑壔偟偰偒偨怉暔奅偲戝偒偔堎側傞丅怉暔偼岝崌惉偵傛傝丄恎嬤偵偁傞悈偲擇巁壔扽慺偐傜帺傜偺懱傪偮偔傝丄惗偒傞堊偺僄僱儖僊乕傪帺暘偱惗嶻偡傞偙偲偑偱偒偨丅怉暔偼摦偔昁梫偑側偐偭偨偺偱丄摦偔堊偺婍姱傗徚壔丒攔煏傗屇媧偺堊偺婍姱傪帩偨側偔偰椙偐偭偨偺偱偁傞丅偦偺偨傔丄怉暔偼嵶朎傪儗儞僈傪愊傓傛偆偵愙拝偝偣偰嶌傞偩偗偺娙扨側峔憿偺懱偱帠懌傝偨丅偲偙傠偑丄怉暔偼堚揱巕偺懡條惈傪幚尰偡傞偨傔偺懠偺屌懱偲偺惗怋偵嬯楯偡傞帠偵側偭偨丅怉暔偼摦偗側偄懱偱壗偲偐懡條側堚揱巕傪幚尰偟傛偆偲偟偰條乆側惗怋朄傪帋傒丄偦偺夵椙偱恑壔偟偰偒偨乯
丂嵟弶偺摦暔偼扨嵶朎惗暔偑扨偵婑傝揧偄晅拝偟偰夠傑傝忬偵側偭偰惗妶偡傞孮懱摦暔偱<柍泱梩摦暔>偲偄傢傟傞丅奀柸摦暔偑偦偺戙昞丅
丂偦偺偆偪夠傑傝忬偺嵶朎偵暘嬈偑惗偠偰撪懁偺徚壔傪巌傞傕偺偲懱昞傪峔惉偡傞傕偺偵暘偐傟偰戃忬偺懡嵶朎惗暔偑惗傑傟偨丅戃偺岥偐傜妉暔傪庢傝擖傟丄偦偟偰徚壔偟偨巆傝偐偡傪偦傟偐傜攔弌偟偨丅偙偺抜奒偱偼傑偩丄岥偲汨栧偑嫟捠側偺偩偑丄戃偺奜懁偲撪懁偺擇憌偺戃偑偱偒傞偺偱<擇泱梩摦暔>偲偄傢傟傞丅巋朎摦暔丒桳孂摦暔側偳偑偁傞丅
丂師側傞恑壔偼戃偑摏忬偵側傝丄妉暔傪庢傝擖傟丄徚壔丄媧廂偟丄巆傝偐偡傪攔弌偡傞偙偲偑棳傟嶌嬈偱弴彉傛偔擻棪揑偵峴偊傞傛偆偵偡傞偙偲偩偭偨丅摏偵偡傞偵偼戃偺岥偺斀懳懁偵寠傪奐偗偹偽側傜側偄偑丄僷僀僾偺巊偄曽偱摦暔奅偼戝偒偔擇偮偵暘偐傟偨丅偡側傢偪丄嵟弶偺岥乮尨岥乯偑偦偺傑傑岥偵側傝丄偦偟偰娧捠偟偨寠偑汨栧偵側傞媽岥乮愭岥乯摦暔偲丄尨岥偑汨栧偵丄娧捠偟偰屻偐傜偱偒偨寠偑岥偵側傞怴岥乮屻岥乯摦暔偱偁傞丅偙偺抜奒偵側傞偲徚壔婍姱傗屇媧婍姱傪峔惉偡傞撪泱梩丄昞旂傗恄宱宯傪峔惉偡傞奜泱梩偲丄偦偺娫偱嬝擏傗惗怋婍姱丄弞娐婍姱丄攔煏婍姱傪峔惉偡傞拞泱梩偺嶰晹峔惉偲側傝<嶰泱梩摦暔>偲偄傢傟傞丅
丂媽岥乮愭岥乯摦暔偼丄嵟嬤偺儕儃僜乕儉俼俶俙偺堚揱巕夝愅偐傜偙傟傑偱偺恀懱峯摦暔丒尨懱峯摦暔偺暘椶偼尒偣偐偗偺傕偺偱丄慄宍摦暔丄愡懌摦暔偺扙旂摦暔僌儖乕僾偲椫宍摦暔丄娐宍摦暔丄擃懱摦暔偺姤椫摦暔僌儖乕僾偵暘偗傞傎偆偑揔愗偱偁傞帠偑夝偭偰偒偨丅偙偺拞偱愡懌摦暔栧亅崺拵峧偼抧媴忋偱嵟傕惉岟偟偨摦暔偺峧偱偁傞丅
丂怴岥乮屻岥乯摦暔偺尨巒揑側僌儖乕僾偑灆旂摦暔偱偁傞偑丄傗偑偰懱偺拞怱偵塣摦帪偺恈偲側傞愐嶕偑惗偠傞丅偙偺抜奒傪尨嶕摦暔偲偄偆丅偦偺愐嶕偼偝傜偵婡擻揑側愐捙偵抲偒姺偊傜傟偰嵟傕桪傟偨塣摦擻椡傪帩偭偨愐捙摦暔偑惗傑傟傞丅偙偺側偐偵嫑椶丄椉惗椶丄唳拵椶丄榢椶丄捁椶丄歁擕椶偑娷傑傟傞丅
丂変乆恖椶偼堦墳丂摦暔奅仺愐捙摦暔栧仺歁擕峧仺楈挿栚乮僒儖栚乯仺恀墡垷栚仺嫹旲壓栚仺僸僩壢仺儂儌懏仺僒僺僄儞僗庬丂偲側傞偑丄忬嫷偵傛傝奒掤偺暘偗曽偼曄傢偭偰偔傞丅
丂壓恾偺媽岥摦暔丒怴岥摦暔偺戝暘椶孮偼丄嵟嬤偺暘巕惗暔妛偵傛傞堚揱巕夝愅偺寢壥偱傕嫮偔巟帩偝傟偰偄傞丅偟偐偟嵶晹偵偮偄偰偼曄峏傪敆傞帠幚偑師乆偲柧傜偐偵側傝偮偮偁傞丅偦偺偨傔壓恾偺宯摑庽偼夵掶偑昁梫偱偁傞丅
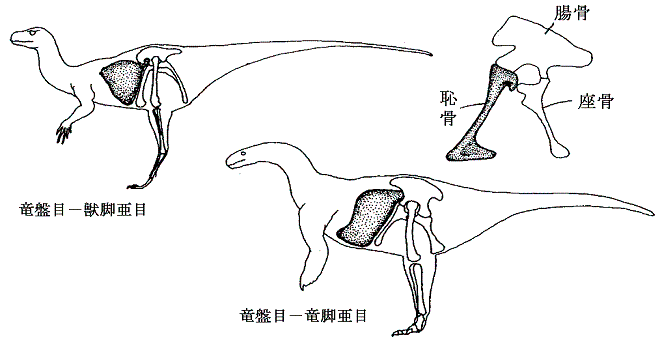
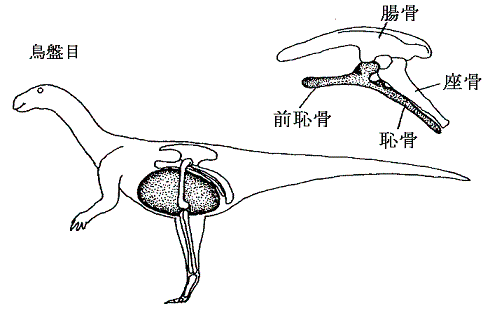
丂廬棃丄摦暔偺懱傪嶌傞懱峯偺宍懺偼丄摦暔偺暋嶨壔丄戝宆壔偱廳梫側栶妱傪墘偠偨偲峫偊傜傟偰丄摦暔偺宯摑暘椶偺戝偒側嫆傝強偲側偭偰偄偨丅偮傑傝丄柍懱峯摦暔乮奜泱梩偲撪泱梩偺娫偵偒傢偩偭偨寗娫偼側偄乯仺婾懱峯摦暔乮奜泱梩偲撪泱梩偺娫偵婍姱偑娷傑傟傞嬻娫乮懱峯乯偑惗偠傞乯仺恀懱峯摦暔乮拞泱梩偱撪挘傝偝傟偨懱峯偑條乆側婍姱傪廂擺乯偲暋嶨壔偟偰恑壔偟偰偒偨偲峫偊傜傟偰偄偨丅偦偺偨傔柍懱峯摦暔偱偁傞滸宍摦暔栧乮僾儔僫儕傾側偳乯傗婾懱峯摦暔偱偁傞慄宍摦暔栧乮僙儞僠儏僂側偳乯偼丄媽岥丒怴岥摦暔偺暘婒傛傝慜偺恑壔偺憗偄抜奒偱暘婒偟偨尨巒揑側摦暔偲峫偊傜傟偰偄偨丅
丂偟偐偟嵟嬤偺儕儃僜乕儉俼俶俙偺堚揱巕夝愅乮傾僌僀僫儖僪傜乯偺寢壥偼丄偦偺條側峫偊曽偑惉傝棫偨側偄偙偲傪帵偟偰偄傞丅堚揱巕夝愅偐傜摼傜傟偨偦傟傜偺摦暔偺暘婒擭戙偼丄媽岥摦暔偺恀懱峯摦暔偑惗偠偨屻偵婲偙偭偨帠傪帵偟偰偄傞丅偦偺偨傔懱峯偺條幃偼丄宯摑暘椶傪寛掕偯偗傞杮幙揑側宍懺揑摿挜偲偼尵偊側偔側偭偨丅屻偐傜梈捠柍奦偵偄偐傛偆偵偱傕曄壔偡傞傕偺偱丄柍懱峯乮滸宍摦暔乯傗婾懱峯乮慄宍摦暔乯傕屻偐傜惗偠偨戅壔揑側宍懺偐傕偟傟側偄丅
丂
丂奀偐傜棨偵忋偑偭偰恑壔偟巒傔偨愐捙摦暔乮梤枌椶乯偼丄崱擔懁摢崪偵奐偄偨寠偺忬嫷偱暘椶偝傟傞丅妠傪暵偠傞偲偒偼杍偺暻偺壓曽偵偁傞懁摢嬝偑廂弅偟偰奜懁偵朿傟傞丅偦偺偲偒朿傟偁偑偭偨嬝擏傪帯傔傞寠偑偁傞曽偑傛傝戝偒偔朿挘偡傞帠偑偱偒傞丅偦偆偡傞偲媡偵懁摢嬝偑怢挿偟偨偲偒偼丄傛傝懡偔偺怢挿偑壜擻偵側傝岥傪傛傝戝偒偔奐偔偙偲偑偱偒傞丅偦偺偨傔懁摢晹偵奐偄偨寠偼妠偺婡擻傪旘桇揑偵夵椙偟偨偲峫偊傜傟偰偄傞丅乮傾儖僼儗僢僪丒儘乕儅乕乯

丂懁摢晹偵奐偄偨寠偺偨傔偵丄慜摢晹偑屻摢晹偲傾乕僠乮媩乯偱宷偑傟峔憿偵側偭偨丅偦偺偨傔傾乕僠偺側偄柍媩椶丄寠偑摢崪偺崅偄埵抲偵偁傞偨傔傾乕僠偺暆偑峀偔側偭偨峀媩椶丄堦偮偺傾乕僠偺扨媩椶丄擇偮偺傾乕僠傪帩偮憃媩椶偵暘椶偝傟傞丅
扨媩椶偼妉暔傪廳偄帟偲嫮椡側妠嬝偱偐傒嵱偔愴棯傪慖戰偟偨丅偦偺偨傔廳偄摢晹偑昁梫偵側傝巐偮懌曕峴傪懕偗偨丅偙傟偺巕懛偑崱擔偺歁擕椶丅
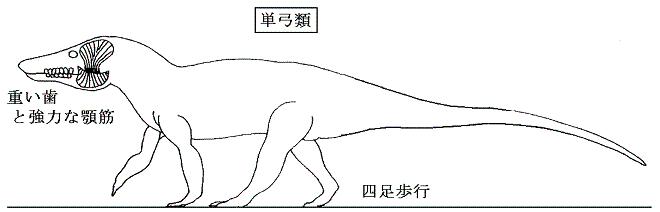
憛帟椶乮憃媩椶乯偼擇杮懌偺巔惃偺偨傔摢崪偑寉偔側偔偰偼側傜側偐偭偨偺偱嵒偺偆偑恑壔偟偨丅偦偺拞偱怘暔偼愇偵傛偭偰偡傝偮傇偝傟僪儘僪儘偵側傞丅嵒偺偆偼憛帟椶偺懱偺廳怱偵嬤偄強偵崌偭偨偺偱丄戝偒側摢傪帩偮扨媩椶偵斾傋偰偼傞偐偵憱傝傗偡偐偭偨丅憛帟椶偺摢晹偼妉暔傪偐傒嵱偔傛傝傕曔傜偊偰嶦偡偙偲偵巊傢傟傞婍姱偩偭偨丅拞惗戙偵偼偄傢備傞嫲棾椶偲偟偰戝斏怋偟偨偑偦偺傎偲傫偳偑拞惗戙枛偵愨柵偟偰偟傑偭偨丅偙偺宯摑偱崱擔惗偒巆偭偰偄傞偺偑唳拵椶丒榢椶丒捁椶偱偁傞丅
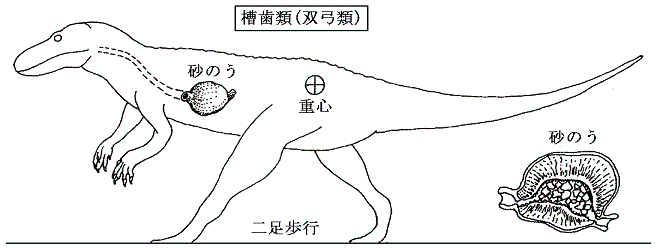
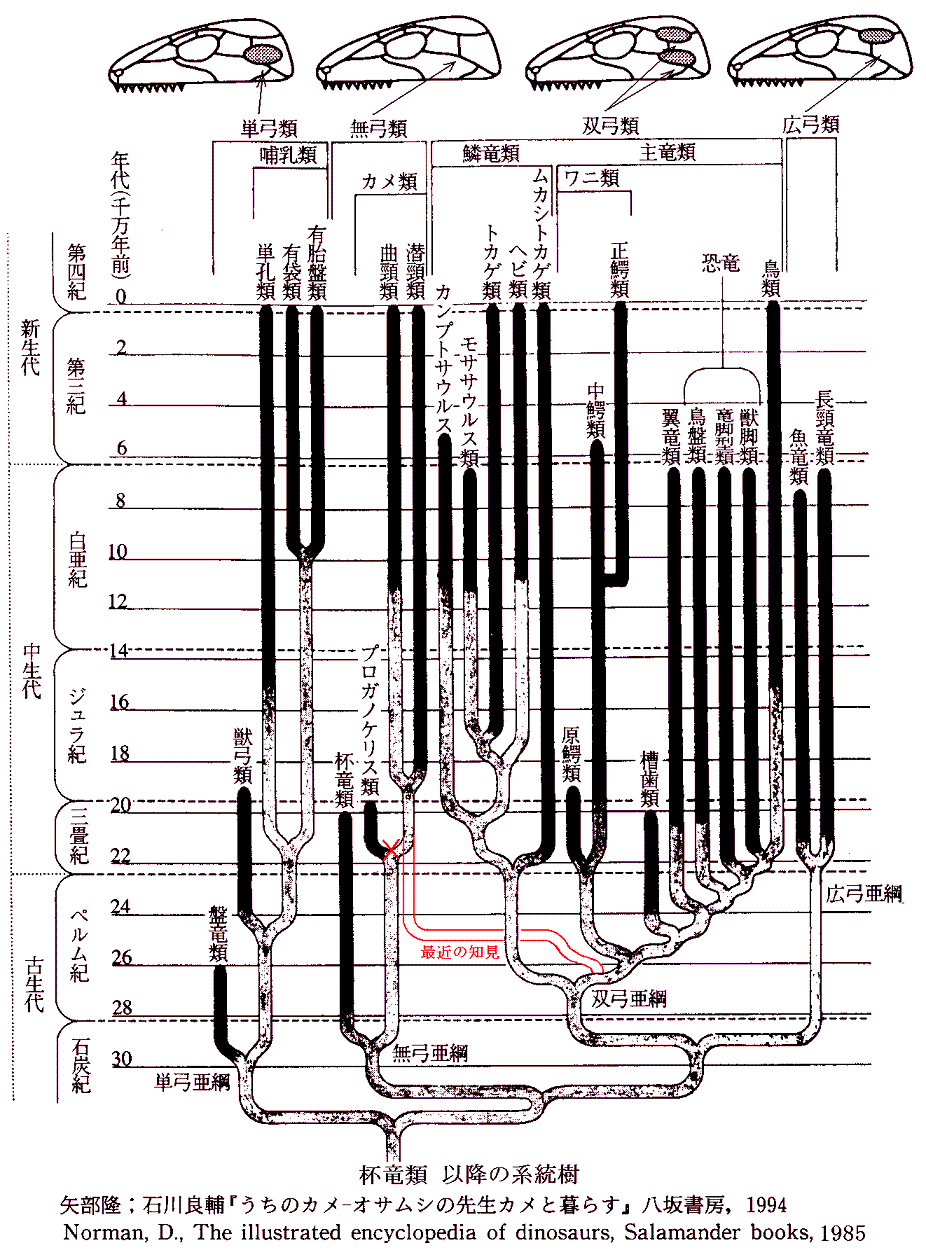
丂忋恾偼廬棃偺宯摑庽偱偁傞丅梤枌椶偺拞偱嵟弶偵懠偐傜暘偐傟偨偺偼扨媩椶乮歁擕椶乯偱偁傝丄偙偺偙偲偼宍懺妛揑偵傕丄暘巕宯摑妛偐傜傕巟帩偝傟偰偄傞丅偟偐偟丄巆傝偺宯摑偺側偐偱丄師偵暘婒偟偨嵟傕屆偄宯摑偲峫偊傜傟偰偄偨柍媩椶乮僇儊椶乯偺埵抲偵偮偄偰暘巕堚揱妛偐傜曄峏傪敆傜傟偰偄傞丅儈僩僐儞僪儕傾俢俶俙乮儊僀儎乕仌僓儖僪僀僫丄惣揷仌孎戲乯傗妀俢俶俙乮媨揷乯偺夝愅偐傜扨媩椶偺暘婒偼僿價傗僩僇僎偺唳拵椶傛傝傕屻偱丄憃媩椶偺僩儕丄儚僯偵嬤偄宯摑偱偁傞偙偲偑夝偭偰偒偨丅偦偺偨傔丄柍媩椶偼恑壔偺夁掱偱屻偐傜懁摢寠傪幐偭偨僌儖乕僾偱偁傞偐傕偟傟側偄丅
丂
丂偄傢備傞嫲棾椶偼崪斦偺宍偱戝偒偔擇偮偵暘椶偝傟傞丅徚壔婍姱偑抪崪偺慜偵偁傞棾斦椶偲抪崪偺壓偵偁傞捁斦椶偱偁傞丅
丂擇懌曕峴偺擏怘惈嫲棾乮憃媩垷峧亅棾斦栚亅廱媟垷栚乯偼斾妑揑彫偝側徚壔婍姱偑抪崪偺忋偵偁偭偨丅偦傟偼廳怱偺慜偵偁傝擇懌曕峴偱晀彿側峴摦偟偨丅憪怘惈嫲棾乮憃媩垷峧亅棾斦栚亅棾媟垷栚乯偼嫄戝側徚壔婍姱傪昁梫偲偟偨丅偦偺徚壔婍姱偼廳怱偺慜偵埵抲偡傞偨傔丄偦偺廳検傪慜懌偱巟偊傞偨傔偵巐懌曕峴偲側偭偨丅
捁斦椶乮憃媩垷峧亅捁斦栚乯偱偼嫄戝側徚壔婍姱偵埑敆偝傟偨偙偲偵傛傝丄抪崪傪屻曽偵堏摦偝偣傞曽嶔傪偲偭偨丅偦偺偨傔嫄戝側徚壔婍姱傪崪斦偺壓偺廳怱埵抲偵廂擺偡傞偙偲偑壜擻偵側傝丄擇懌曕峴傪堐帩偡傞傕偺偲巐懌曕峴偺傕偺偑恑壔偟偨丅
丂嫲棾偺墹條僥傿儔僲僒僂儖僗乮俿丒儗僢僋僗乯偼廬棃丄傾儘僒僂儖僗偲摨偠僇儖僲僒僂儖僗椶偵懏偟偰偄傞偲偝傟偰偄偨偑丄嵟嬤偺尋媶偵傛傞偲丄僐僄儖儘僒僂儖僗椶偵懏偡傞傕偺偱丄傛傝尰戙偺捁椶偵嬤偄宯摑偺擏怘嫲棾偱偁傞偙偲偑夝偭偰偒偨丅
乵2017擭4寧5擔捛婰乶
丂忋婰偺崢崪偺宍偵傛傞戝暘椶偺掕愢乮1887擭Harry Govier Seeley偑敪昞乯傪暍偡怴偟偄暘椶偑嵟嬤乮2017擭3寧23擔乯働儞僽儕僢僕戝妛偺尋媶僠乕儉偵傛傝敪昞傟偝偨丅
乬New study shakes the roots of the dinosaur family tree乭
Matthew G.Baron, etal., 乬A new hypothesis of dinosaur relationships and early dinosaur evolution乭, Nature, 545, p501乣506,(23,March,2017)
丂
丂偙偺怴愢偵傛傞偲丄T丒儗僢僋僗側偳屻偵捁椶偵恑壔偟偨偲偝傟傞嫲棾偼廬棃丄乲棾斦椶乴偺壓埵暘椶偱偁傞乹廱媟椶乺偵暘椶偝傟偰偒偨偑丄T丒儗僢僋僗側偳偺乹廱媟椶乺偼乲棾斦椶乴偐傜奜偝傟丄僗僥僑僒僂儖僗側偳偺乲捁斦椶乴偲摨偠怴偟偄僌儖乕僾偵暘椶偝傟傞丅
丂偵傢偐偵偼怣偠偑偨偄榖偱偡偑丄傕偟偙傟偑杮摉側傜暘椶宯摑庽偺戝曄壔偱偡偹丅
丂
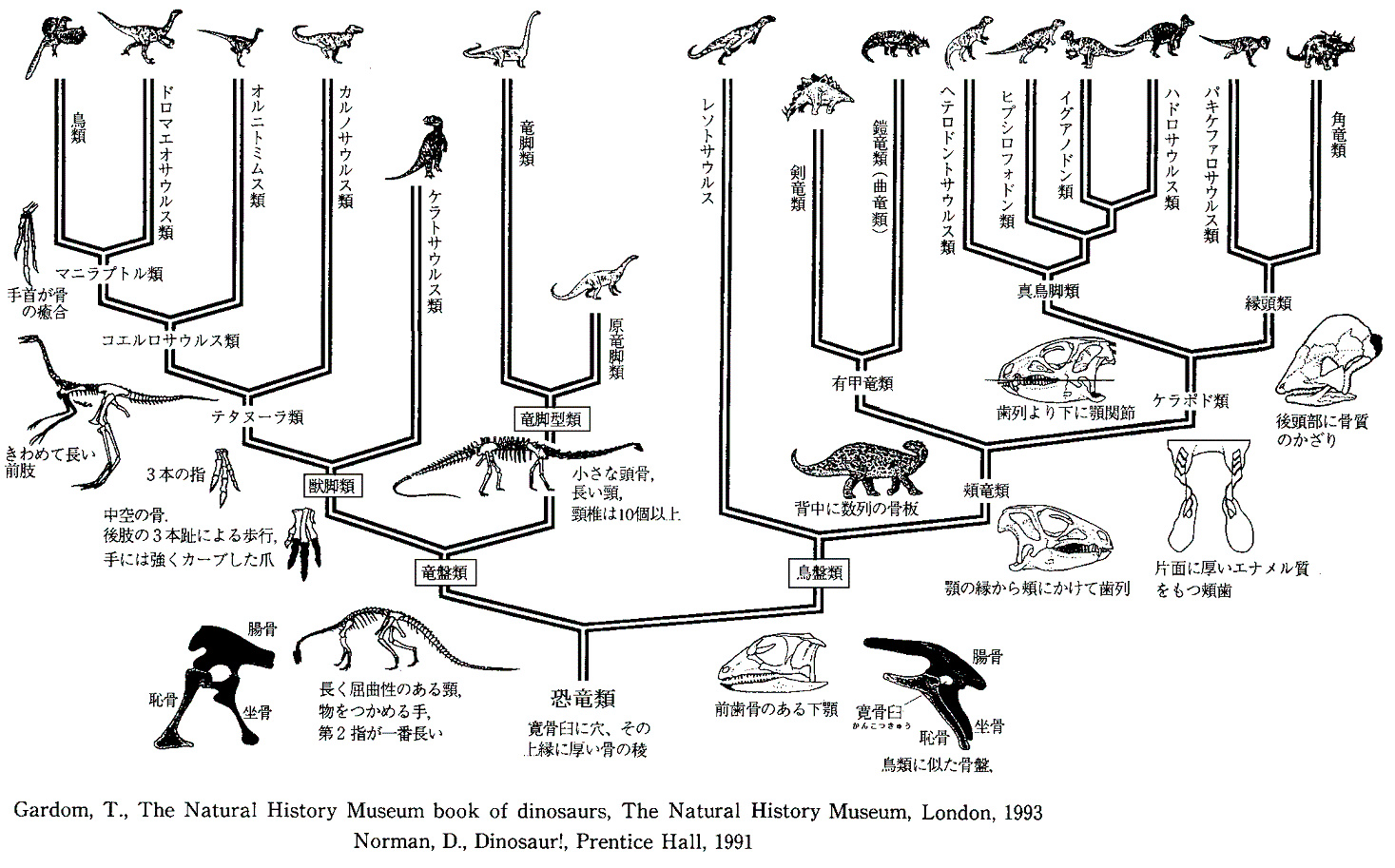
丂摦暔恑壔偺夁掱偱婲偙偭偨嶰偮偺廳梫側堚揱揑曄堎乮侾丏媽岥丄怴岥偺暘婒丄俀丏懁摢崪偵惗偠偨寠偺嵼傞柍偟丄俁丏崪斦偺曄壔乯傪徯夘偟偨丅偙傟傜偺堚揱揑曄堎偼庬撪偵屌掕偝傟挿偔宲彸偝傟偰偒偨傕偺偱暘椶妛忋嵟傕婎杮揑側曄堎偱偁傞偲峫偊傜傟偰偄傞丅偟偐偟丄偦傟傜偺曄堎偑揔幰惗懚偺帺慠慖戰偵懳偟偰桳棙偐偳偆偐偼壗偲傕尵偊側偄丅暘婒偟偨暿乆偺宯摑傕師悽戙偵揱偊傜傟偰懚懕偟偨偺偩偐傜丅
丂忋偵弎傋偨條偵丄暘巕堚揱妛偵傛傞嵟嬤偺抦尒偺嫵偊傞偲偙傠偵傛傞偲丄宍懺妛揑側婡擻曄壔偼恑壔偦偺傕偺偵偼傎偲傫偳塭嬁偟側偄傛偆偱偁傞丅偙偙偺偲偙傠偼埲壓偺傛偆偵傑偲傔傜傟傞傛偆偵巚偆丅
寢嬊偺偲偙傠丄惗暔偼忋偵弎傋偨係偮偺抜奒偺係偮偺尨棟偵廬偭偰恑壔偟偰棃偨傛偆偵尒偊傞丅